

Plants and animals are shifting their ranges in response to recent climate change4, 5 and the climate predicted for the twenty-first century will further increase pressure on species to shift their geographical distributions6. In many terrestrial environments, however, human-modified landscapes offer little space for the establishment of new populations and impede the redistribution of species7, 8. Indeed, empirical evidence suggests that species frequently prefer high-quality habitats as offered by protected areas when expanding their ranges9. As a consequence, increasing the quality, quantity and connectivity of habitats appears pivotal for mitigating negative impacts of climate change on biodiversity3, 10.
Three important approaches for the spatial allocation of habitat-based climate change adaptation measures are: increasing the conservation value of the existing protected area network—that is, improving habitat quality and/or management efficiency within protected areas11; establishing high-quality habitat corridors to enhance connectivity among protected areas12; and improving the habitat quality and permeability of the ‘average’ landscape matrix, for example, by establishing interspersed high-quality habitat patches such as low-intensity grasslands, hedgerows, marginal field strips or (semi-)natural forest patches13. In Europe, each of these conservation strategies has recently been integrated into spatial planning and land use policies such as the European Habitats Directive14, the European Union Green Infrastructure strategy15, the European Union agriculture policy, and ecosystem restoration goals16. However, whether these strategies mitigate climate-change-driven range loss of species remains little tested so far17.
Here, we used coupled niche-demographic modelling18 (see Supplementary Fig. 1) to assess regional range loss and extinction risk of 51 species from three taxonomic groups (vascular plants, butterflies, grasshoppers) under current climatic conditions and two climate change scenarios (B2—mild scenario, A1B—moderately pronounced scenario) in Central Europe (Austria, Switzerland, Liechtenstein, Northern Italy and southern Germany, Supplementary Fig. 2). We thereby compared a ‘business-as-usual’ setting (that is, current spatial habitat patterns remain unchanged) with three conservation strategies: improving protected area management (P), establishing corridors (C) and improving the landscape matrix (M). The 51 species represent a wide range of ecological profiles (Supplementary Tables 1–3) and have their current elevational distribution centred either in the colline to montane (henceforth termed lowland species) or the subalpine to alpine belt (alpine species; 7–10 species from each elevational group per taxon depending on model quality—see Supplementary Methods). Conservation strategies were implemented by modifying a gridded habitat distribution map (spatial grain 100 × 100 m; in the following, cells of this grid are called ‘sites’) of the study region19—that is, by converting a proportion of intensively used habitats (agricultural fields, nutrient-rich grasslands, conifer plantations) to extensively used semi-natural ones (for example, dry grasslands, wet grasslands, broadleaved forests). We applied three effort levels: low (that is, 1% of sites being converted), medium (3%) and high (5%). Conservation strategies differed in the spatial arrangement of habitat conversion. In P (protected areas) and C (corridors) conversion was restricted to protected areas or to protected areas and connecting corridors, respectively (Supplementary Fig. 2). In M (matrix), converted habitats were randomly distributed across the landscape. We evaluated the effectiveness of these strategies as compared to the business-as-usual strategy by simulating changes in range size of the study species until the year 2090. To account for uncertainties in the estimation of the species’ demographic and dispersal rates, all simulations were run using two sets of parameters, one at the upper and one at the lower end of documented parameter ranges (Supplementary Tables 2 and 3).
Under unchanged climatic conditions and without any conservation measures, our simulations resulted in approximately constant range sizes of all model species until the end of the century (Supplementary Fig. 3). With climate warming, however, about one in five species was predicted to go regionally extinct under both the mild (1 alpine/0 lowland, 1/1 and 3/3 species for plants, butterflies and grasshoppers, respectively) and moderately pronounced scenario (1/1, 1/0 and 3/4 species for plants, butterflies and grasshoppers, respectively). Modelled conservation strategies and efforts had no effect on the fate of these species: none of them was rescued, even under the highest effort levels.
For those species not going extinct (9 alpine/9 lowland plants, 8/6 butterflies and 5/5 grasshoppers under B2 and 9/8, 8/7 and 5/4 under A1B), climate change triggered a reduction in range size which was, on average, similarly pronounced under the A1B (mean: 58% and 61% under the high and low demographic and dispersal parameter sets, respectively) and the B2 scenario (52%/62%), but much stronger for lowland than for alpine species (Table 1). Conservation strategies were more effective in reducing range loss than in avoiding extinction with, again, pronounced differences between lowland and alpine species as well as between taxonomic groups (Fig. 1 and Table 2 and Supplementary Figs 4 and 5). Efficiency was highest for grasshoppers, less marked in the case of lowland butterflies and lowland plants, and marginal in the case of alpine butterflies and alpine plants. In addition, positive effects of conservation strategies were substantially weaker for forest species than for grassland species (Supplementary Fig. 6). Among grassland species, those of dry and extensive grasslands responded more strongly to conservation strategies than those of wet grasslands.
Effects of climate change were calculated as log (range size at 2090/range size at 2010) and effects of conservation strategies as log (range size in 2090 applying a conservation strategy/range size in 2090 under the business-as-usual strategy), respectively. Thus negative values denote range loss, and positive values denote range gain. Results of simulations assuming current climatic conditions to be stable during the twenty-first century are indicated by a grey background, while bars in front of a white background are results of simulations which assume climate change according to the A1B scenario. Results are similar for mild (B2) climate change (Supplementary Fig. 4). Conservation strategies were applied by converting intensively used habitats into habitats of high conservation value within protected areas (P; red label text), within corridors connecting protected areas (C; blue) and within the landscape matrix (M; black). For each conservation strategy, three levels of effort were analysed: low (that is, 1% of sites were converted; represented by P1, C1, M1), medium (3%; represented by P3, C3, M3) and high (5%; represented by P5, C5, M5). Results were averaged over five simulation runs. Species predicted to be committed to extinction until 2090 were excluded from the analysis. Demographic rates were assumed to be at the upper end of the documented range. Number of species: 9 alpine/8 lowland, 8/7 and 5/4 for plants, butterflies and grasshoppers, respectively (see Supplementary Table 1). Results of simulations using the lower parameter set are given in Supplementary Fig. 5.
Under low demographic and dispersal parameters, increasing conservation efforts had only marginal to weak consequences for simulated range loss under climate change (Supplementary Fig. 5). Under high values of demographic and dispersal parameters, by contrast, increasing the effort also increased the impact of conservation measures: for about every fifth lowland grasshopper species, high conservation efforts resulted in late twenty-first-century ranges at least twice as large as under the business-as-usual strategy (Fig. 2). Anyway, for none of the taxonomic groups even the combination of highest level conservation efforts and high parameter choices resulted in full compensation of climate-change-induced average range loss (Fig. 1). Species benefited most from conservation measures under current climatic conditions. Measured effectiveness differed moderately, but statistically significantly, among the three conservation strategies (P > C > M; Table 3). Differences were more significant with low than with high conservation effort (Table 3).
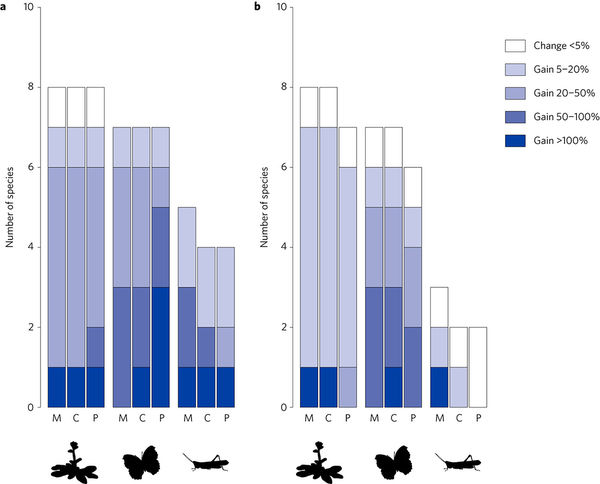
a,b Changes in range size are calculated for the year 2090 by comparing the area simulated to be occupied under a high conservation effort strategy with the area simulated to be occupied under a business-as-usual strategy (that is, no habitat conversion), under either the current climate (a) or moderately pronounced climate change (A1B; b). The proportion of species is given for protected areas (P), corridors (C) and landscape matrix (M) conservation strategies. Results were averaged over five simulation runs. Results assuming mild (B2) climate change are similar to those of pronounced climate change and, hence, not shown.
Taken together, our simulations suggest that regional ranges of all modelled species will decline under a warming climate, and that the efficiency of the tested conservation strategies for halting this decline is limited. However, effects vary considerably among species, with a marked difference among those of low and high elevations. The likely reason is that semi-natural habitats are restricted to remnant patches in Central European lowlands while they still represent the matrix at (sub-) alpine elevations. As a consequence, habitat restoration has ample scope in lowlands but limited potential in high mountain landscapes. In addition, most alpine species are unable to survive in forests. Improving connectivity for their above-treeline habitats would hence require the replacement of subalpine or montane forests, including natural ones, by non-forest habitat types—that is, an increasing intensity of human land use rather than restoring the (semi-)natural forest vegetation of these elevations20.
Even for lowland species, the success of conservation measures is mixed, but improving habitats in protected areas or connecting protected areas via corridors was generally more efficient than randomly distributing high-quality habitats across the landscape. This difference indicates that spatial coherence of suitable habitat is important for species’ survival and range adaptation (Supplementary Fig. 2). Put another way, random placement of habitat patches in an intensively used matrix leaves gaps among potential stepping stones which apparently are often too large for the ‘average’ dispersal capacity of plant or insect species. The spatially condensed habitat conversion under P and C, by contrast, targets those parts of the study region which offer most high-quality patches already today (that is, protected areas) and hence removes dispersal barriers within and between these hotspots of current species distribution.
Lower effectiveness of conservation measures under climate warming is probably due to limited dispersal abilities. An increased temperature forces species to expand their ranges at the front edge of their distribution to track the shifting climate and compensate losses at the rear edge7. The required velocity may easily be beyond species dispersal capacity even in continuous habitats8, a situation far from being reached in our study area even under highest conservation efforts (Supplementary Fig. 2). As a consequence, the additional habitat patches established by conservation measures may be climatically suitable for too short a time to be used as stepping stones during range shifts. Short-lived species such as butterflies and grasshoppers are probably more affected by high climatic velocities because they lack adequate strategies to persist under adverse climates21. An increasing frequency of unfavourable years hence raises their local extinction risk more than those of the mostly long-lived plants in our species set. Grasshoppers, in particular, combine short life cycles with a high degree of ecological specialization. This combination probably makes them highly sensitive to climate effects, but also especially responsive to conservation measures.
We emphasize that the high-level conservation efforts—that is, a conversion of 5% of the study area into high-quality habitat—appears ambitious11. However, even these high efforts would only be able to significantly reduce climate-change-driven regional range loss for a minority of species (Fig. 2). In addition, our business-as-usual strategy—that is, assuming current habitat patterns to remain constant—does not actually presume a worst-case scenario. In fact, land use intensification and expansion of infrastructure have caused severe declines of extensively used habitats in Europe and beyond during the recent decades22. This decline has not yet been halted, neither by the expansion of the protected area network23, nor by newly introduced regional policies such as the European Union Restoration Goals16 or the Green Infrastructure strategy15. Increasing land use demands for biofuel and food production24 additionally suggest that a realistic worst-case scenario might involve further shrinkage of high-quality habitats, and hence even higher vulnerability of species to a warming climate.
Our results are derived from simulations which are necessarily based on simplifying assumptions and coarse parameter estimates, and which certainly neglect important co-determinants of species range dynamics18, 25. It is, for example, possible that even our high dispersal distance estimates are too low for some insect species, as they are mainly based on mark-recapture studies in which long-distance flights tend to be under-represented26. Simulations under the two demographic and dispersal parameter sets (Supplementary Tables 2 and 3) suggest that assuming such higher dispersal distances would have increased the efficiency of management measures for these particular species. However, our main result of considerable range loss under climate warming despite high conservation effort was insensitive to the strong contrast in the two parameter sets that we applied across all model species. It is hence unlikely that assuming still higher dispersal distances for a subset of species would have changed our conclusions qualitatively (Supplementary Fig. 5 and Supplementary Discussion).
Consideration of other modulating factors would have certainly further modified the results quantitatively. In particular, we did not include climate-driven changes to the vegetation in our models. Such changes will probably lag behind climate warming considerably27, but may nevertheless be fast enough to modify local habitat suitability for insect species in the decades to come. Moreover, the spatial grain of our simulations and the way climatic data are downscaled preclude proper accounting for microclimatic refugia which may mitigate range loss, especially in alpine areas. Additional unaccounted factors such as climatic extremes28 or human-mediated dispersal29 can decrease local species persistence or increase their mobility and thus affect range change. In essence, however, the two sets of parameters used span a broad range of possible species’ demography and mobility that should also comprise most effects of these modulating factors on range dynamics.
We conclude that habitat-based measures to mitigate the negative effects of climate warming on species’ range sizes are necessary but that their efficiency is limited. Although additional climate change adaptation measures for species are feasible (such as assisted migration programs), these approaches also face severe constraints because they can realistically be implemented for only few charismatic species. Moreover, they may have unforeseen negative consequences for resident biota30. As a corollary, efforts to mitigate climate effects on biodiversity are urgently needed, but are unlikely to be as effective as halting climate change itself.





